MECANISME DE LA CONTRACTION MUSCULAIRE
Niveau des myofibrilles
Les microfilaments d’actine sont associés à deux autres
protéines :
* la tropomyosine, protéine fibrillaire enroulée en torsade
autour de l’actine, avec un pas égal à celui de la spirale
d’actine
* la troponine, elle-même associée à la tropomyosine.
Elle est formée de 3 sous unités : la troponine C, capable de fixer le
Ca2+, la troponine T capable de fixer la tropomyosine, et la troponine
I qui empêche la fixation de l’actine en présence de Ca2+.
|
En l’absence de Ca2+, la troponine
ne fixe pas de Ca2+, la tropomyosine occupe une position
telle que les sites de fixation de la myosine sur l’actine sont
masqués. A la suite d‘un flux de Ca2+ la troponine
fixe ce Ca2+, ce qui provoque le déclenchement d’une
rotation de la tropomyosine autour de l’actine. Les sites de
fixation de la myosine sur l’actine deviennent libres et accessibles.
|
Intervention du calcium dans l’organisation
des filaments fins

|
Les mouvements des différentes protéines les unes par rapport
aux autres se réalisent selon la chronologie suivante :
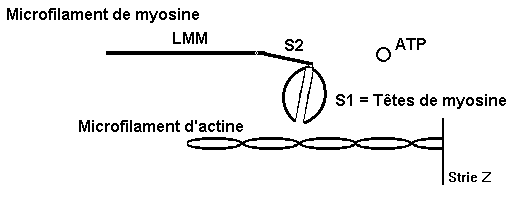
1) 2 têtes de myosine, en s‘associant à une molécule
d’ATP, constituent une tête unique formant un angle de 90° avec le segment
S2 de la molécule de myosine
|
2) Un flux de calcium en provenance du réticulum
sarcoplasmique provoque la rotation de la tropomyosine, ce qui libère
les sites de fixation de la myosine sur l’actine
|
 |
|
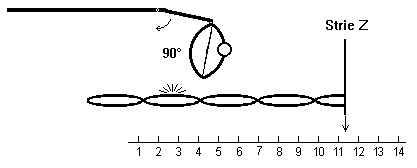
3) l’ensemble ATP-Myosine se fixe sur l’actine
par rotation de la charnière LMM-S2 de la molécule de myosine
|
|
|
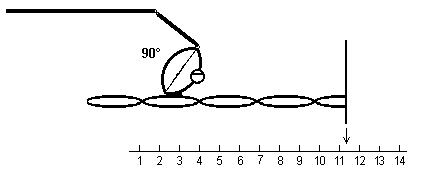
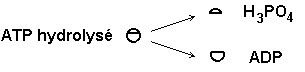
4) L’actine active la fonction ATPase de
la tête de myosine. On obtient l’ensemble actine-myosine-ADP-Pi
|
|
|
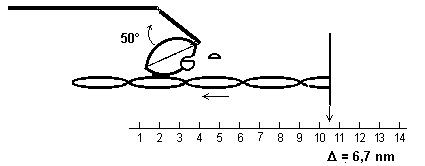
5) La perte de Pi provoque une rotation de la
tête de myosine dont l’angle avec le reste de la molécule de
myosine passe de 90° à 50°, provoquant un glissement de 6,7 nm de
l’actine par rapport à la myosine
|
|
|
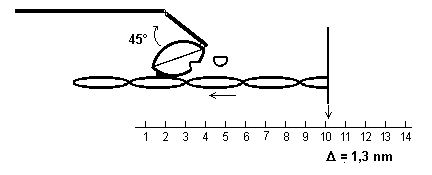
6) La perte de l’ADP provoque un glissement
supplémentaire de 1,3 nm.
Le complexe Actine-myosine est stable.
|
|
La fixation d’une nouvelle molécule d’ATP sur la
myosine provoque une séparation entre actine et ATP-myosine, et en conséquence
le retour à l’état initial relâché.
Il se produit un recul de l’actine par rapport à la myosine
et une rotation inverse de la tête de myosine qui passe de 45 à 90° par rapport
au reste de la molécule de myosine.
En l’absence d’ATP, le complexe actine-mysosine reste
stable. C’est ce qui se produit lors de l’établissement de la rigidité
cadavérique, alors que la production d’ATP est arrêtée.
Si la concentration sarcoplasmique en calcium se maintient,
le mouvement se reproduit. Au contraire, si les pompes à Ca2+ du
réticulum sarcoplasmique provoquent une baisse de la concentration en calcium,
la contraction s’interrompt. Lors d’un déficit en ATP, le fonctionnement
insuffisant des pompes à Ca2+ peut provoquer le phénomène de crampe.
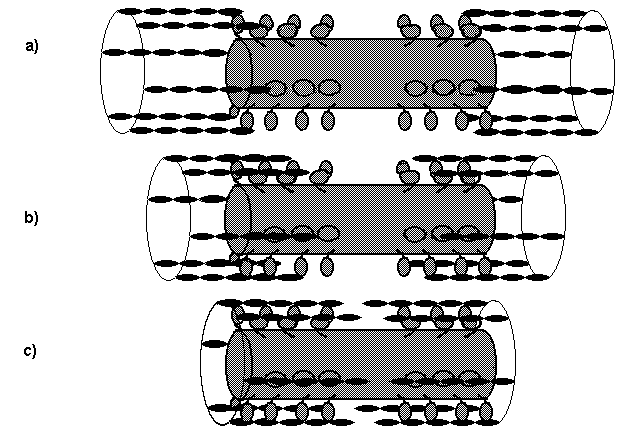
Globalement, le mouvement des têtes de myosine provoquent un
glissement de l’actine qui rapproche les stries Z les unes des autres en
réduisant les zones H et la bande I (Fig. ci-dessous). Un microfilament
peut effectuer 5 cycles par seconde, de façon asynchrone avec les voisines.
Représentation des mouvements relatifs entre filaments fins
et filaments épais dans un sarcomère
a) Distension maximum b) Relachement c) Contraction
maximum